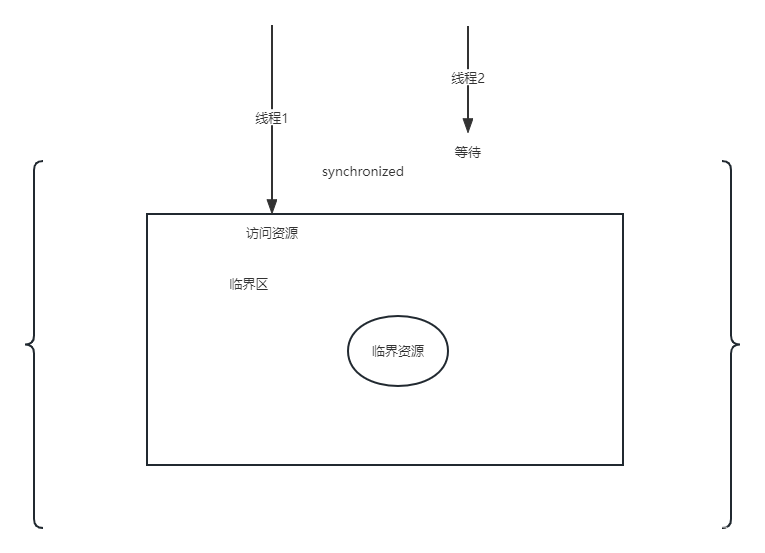
文献介绍

「文献题目」 Inferring neuron-neuron communications from single-cell transcriptomics through NeuronChat
「研究团队」 聂青(加利福尼亚大学欧文分校)
「发表时间」 2023-02-28
「发表期刊」 Nature Communications
「影响因子」 16.6
「DOI」 10.1038/s41467-023-36800-w
摘要
神经通信网络构成了大脑功能的基础。这些通信网络是通过发射配体(如神经递质)来实现的,它们激活受体复合物以促进通信。因此,神经通信基本上取决于转录组。在这里,作者开发了 NeuronChat,一种用于推断、可视化和分析预定义细胞群体之间的神经特异性通信网络的方法和软件包,使用单细胞表达数据。作者整合了人类和小鼠的神经信号传导的手工策划的分子相互作用数据库,并在几个已发表的数据集上对 NeuronChat 进行基准测试,以验证其预测神经连接能力。然后,作者将 NeuronChat 应用于三个不同的神经组织数据集,以说明其在识别神经内部通信网络、揭示不同生物学背景下保守或特定于上下文的相互作用以及预测自闭症谱系障碍患者大脑中通信模式变化方面的功能。最后,作者证明 NeuronChat 可以利用空间转录组学数据来推断和可视化神经特异性细胞-细胞通信。
研究结果
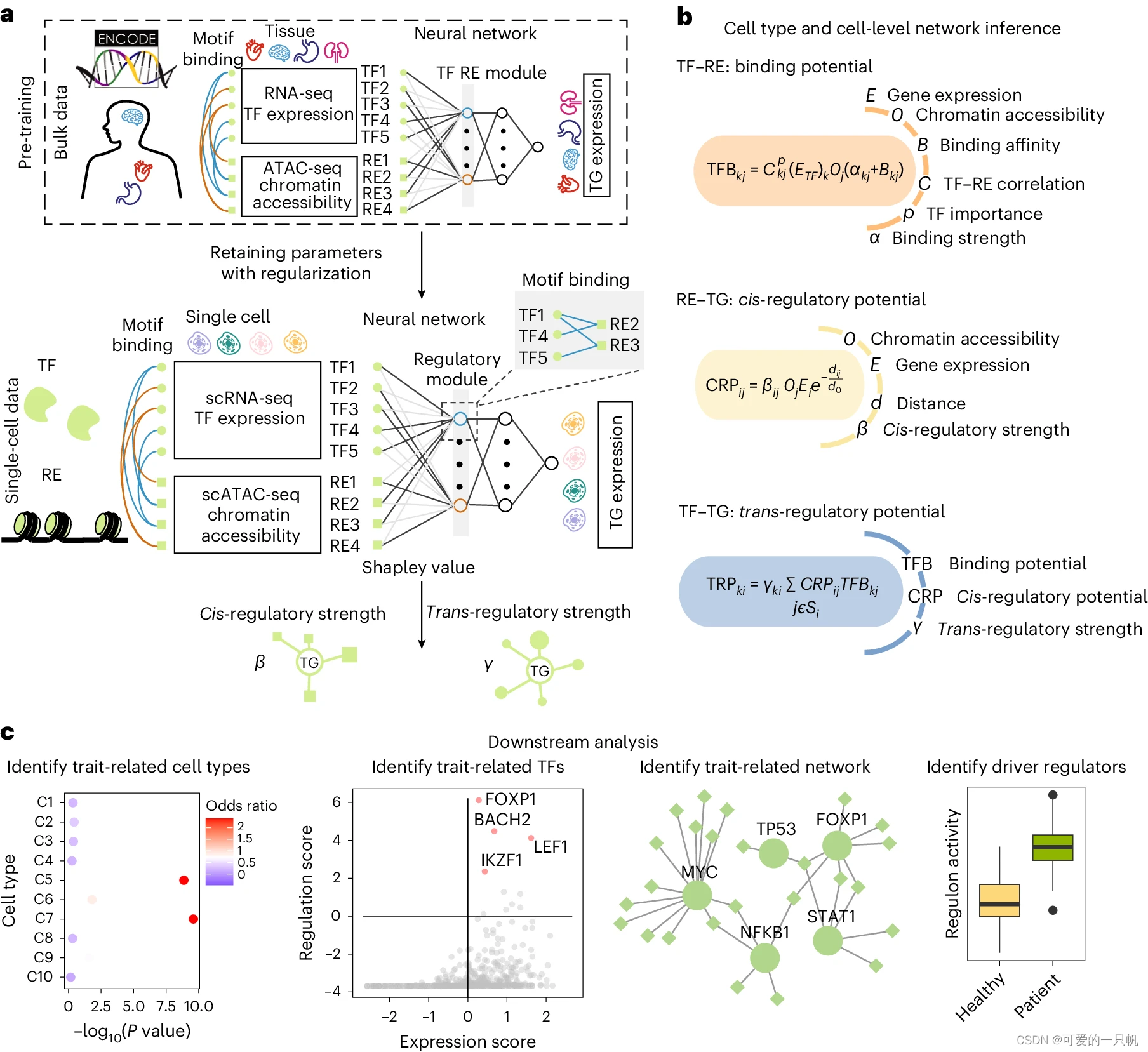
1. NeuronChat 概述
首先,作者为小鼠和人类创建了一个神经特异性的细胞间分子相互作用数据库,名为 NeuronChatDB(Fig. 1a)。每一种相互作用都包含一个 配体(ligand) 和一个同源 靶标(target) 以及与其 合成(synthesis) 和 囊泡运输(vesicular transport) 相关的蛋白质编码基因。ligands 包括小分子神经递质(neurotransmitters)、神经肽(neuropeptides)、间隙连接蛋白(gap junction proteins)、气体递质(gasotransmitters)和突触粘附分子(synaptic adhesion molecules),而 targets 通常但不限于受体。例如,神经递质的靶蛋白也可以是摄取转运蛋白或失活酶;间隙连接蛋白的靶蛋白是其他兼容的间隙连接蛋白。对于非肽类神经递质,条目中包含相应的合成酶和/或囊泡转运蛋白;对于包含多个不同亚基的异聚受体,相应的亚基被整理成具有相同配体的不同条目。在总共 373 个配体-靶点相互作用对条目中,分别有 221、73、39、16、24 个与小分子神经递质、神经肽、间隙连接蛋白、气体递质和突触粘附分子相关的条目。

a. NeuronChat database 概述。NeuronChat database 包括 chemical synapse(化学突触)、electrical synapse(电突触)、synaptic adhesion(突触粘附)所需的 ligand-target pairs(配体-目标对)(左图)。NeuronChat database 共收集了人类和小鼠的 373 对 ligand-target,根据配体的类型分为 5 个类别(中图)。相互作用对列表包括配体、靶标和对它们有贡献的基因(右图)。值得注意的是,参与配体的基因根据其不同的生物学功能,如合成(synthesis)或囊泡运输(vesicular transport),被分为不同的组(用颜色表示)。
b. 说明 NeuronChat 计算模型的示意图。通讯强度表征了发送细胞群中配体发射所需基因的协调表达,以及接收细胞群中目标基因的表达。通信链路的统计显着性由排列检验确定(* 和 ns 分别代表显着和不显着)。仅重要链路保留在输出通信强度矩阵中,而不重要链路的值设置为零。详细信息请参见方法。
c. NeuronChat 的一些功能:细胞间通信网络的可视化与分析,不同生物背景间进行系统的比较,空间转录组学的多层可视化。
其次,作者基于预定义细胞群相互作用分子的坐标表达,构建了一个计算模型,将表达数据与细胞间通讯概率联系起来(Fig. 1b)。NeuronChat 的输入数据是一个 标准化的 cell-by-gene 计数矩阵,带有细胞的 group 注释。对于 NeuronChatDB 中的每个细胞间相互作用对,首先按 cell group 对所有相关基因的表达水平进行平均,基于此估计每个 cell group 的 ligand 和 target 的丰度。对于非肽神经递质,首先将有助于 ligand 发射的基因分类为不同的生物功能组(例如,合成(synthesis)和囊泡运输(vesicular transport)),然后通过在不同组之间应用 AND 逻辑(即几何平均值)来建模丰度。基因,同时在同一组内的冗余基因之间应用 OR 逻辑(即算术平均值);对于其他配体和所有靶标,丰度计算为平均表达。然后,将两组之间的细胞间通讯强度设置为一个 cell group 的 ligand 丰度与另一 cell group 的 target 丰度的乘积。重要的通信可以通过排列测试来确定,其中随机排列 cell group 标签并重新计算通信强度。因此,对于每次相互作用,可以构建细胞间通信网络,即由相互作用的 cell groups 之间的重要链接组成的加权有向图。通过总结与四种不同聚合方法进行单独交互的所有通信网络,可以进一步获得聚合通信网络。
第三,NeuronChat 提供了不同的方法来可视化和分析推断的细胞间通信网络(Fig. 1c)。圆形图、弦图和热图可用于可视化细胞组之间的通信强度。NeuronChat 还可以对推断的通信网络进行定量分析,以识别信号模式并对交互进行分类。对于来自不同生物背景的多个数据集,NeuronChat 可以进行系统比较并识别保守的和特定背景的 ligand-target 相互作用对。对于空间转录组数据,NeuronChat 可以将细胞空间定位纳入细胞间通讯的推理中,并提供空间细胞间通讯的多层可视化。
2. NeuronChat 基准测试
为了研究 NeuronChat 预测细胞间通信的能力,作者首先将预测的通信网络与实验确定的基准进行比较。研究了两个案例:(1)小鼠大脑中初级视觉皮层(VISp)的投射网络,以及(2)小鼠大脑中前外侧运动皮层(ALM)的投射网络。VISp 和 ALM 的兴奋性神经元与其皮质目标区域的连接是使用单突触逆行标记识别的,其中病毒示踪剂被注射到目标区域并通过逆行轴突运输向突触前神经元移动,而不会进一步扩散到间接接触的细胞,从而允许直接神经连接的识别。通过使用细胞类型注释对逆行标记神经元进行分组,获得了粗粒度投射网络,该网络由从 VISp 和 ALM 中的兴奋性神经元类型到其皮质目标区域的定向链接组成(Fig. 2a for VISp and Fig. 2f for ALM) ,然后用于后续基准测试。VIsp、ALM 及其目标区域的 scRNA-seq 数据收集自两篇已发表的论文。使用的数据包括 VIsp 的 7 个亚类(L2/3 IT、L4、L5 IT、L5 PT、L6 CT、L6 IT 和 L6b)的 6,785 个谷氨酸能细胞,以及来自三个皮质目标区域(ACA, RSP and contralateral VISp); ALM 的 5 个亚类(L2/3 IT、L5 IT、L5 PT、L6 CT 和 L6 IT)的 3,883 个谷氨酸能细胞,以及来自六个皮质目标区域(SSs, SSp, RSP, MOp, contralateral ALM and contralateral ORB)。
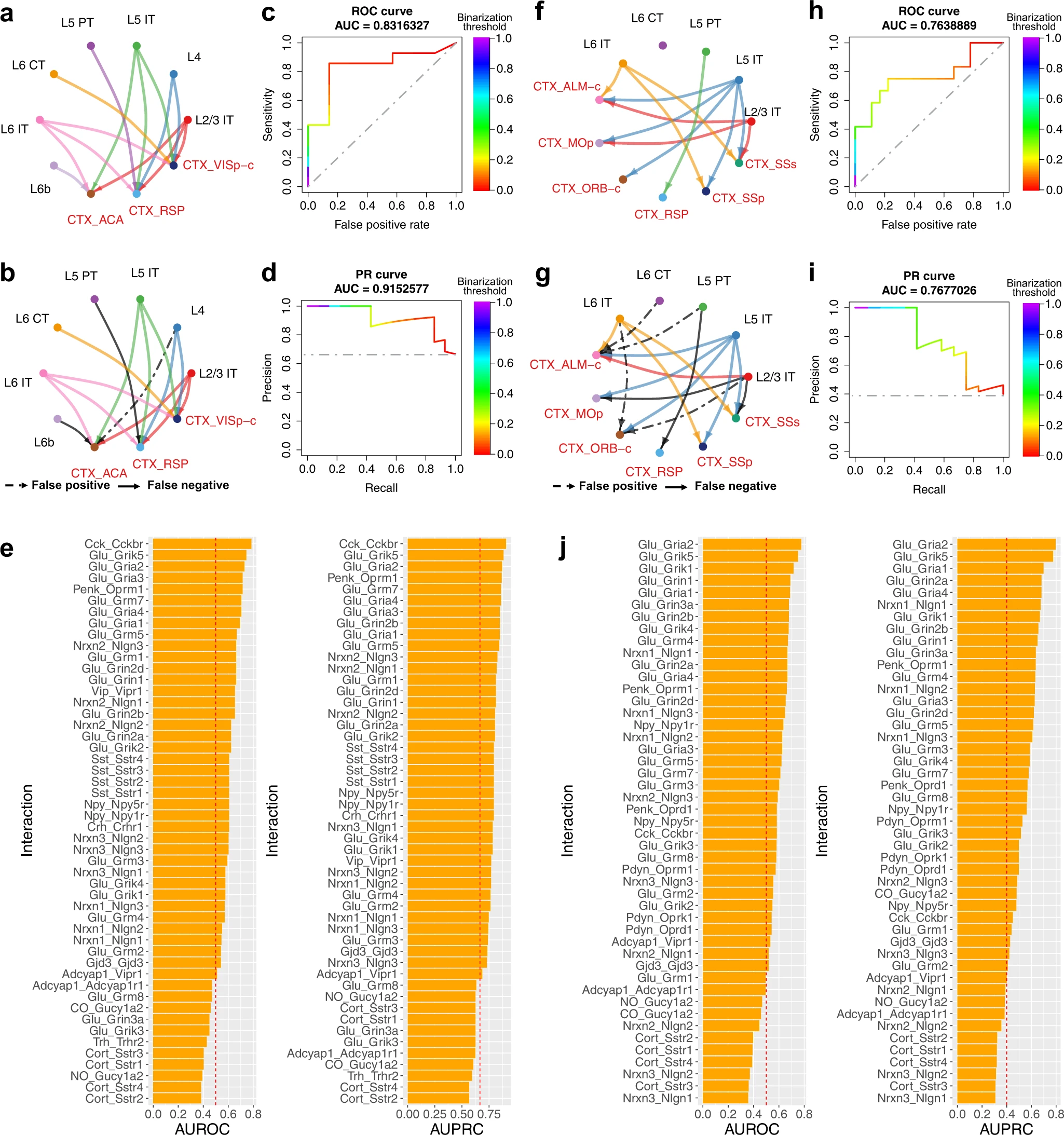
a. 通过逆行标记进行识别的,从 7 种 VISp 细胞类型到其皮层靶区的投射 ground truth。
b. NeuronChat 预测的 VISp 细胞间通信网络。假阳性连接代表了那些被 NeuronChat 预测但没有被逆行标记识别的连接。假阴性连接代表那些被逆行标记识别但被 NeuronChat 预测遗漏的连接。在此图中,二值化的阈值是 0.028(按最大值归一化)。
c. NeuronChat 预测的 VISp 细胞间通信网络的 ROC 曲线,顶部显示了 AUROC 值为 0.8316327。灰色虚线表示 AUROC 为 0.5 的随机分类器。颜色条表示二值化的阈值。
d. NeuronChat 预测的 VISp 细胞间通信网络的 PR 曲线,顶部显示了 AUPRC 值为 0.9152577。灰色虚线表示 AUPRC 为 0.5 的随机分类器。颜色条表示二值化的阈值。
e. 每个 interaction pairs 的 AUROC 和 AUPRC 值,得分越高表明能更好的预测投射。
f-j. 对 ALM 进行的相同分析。
通过使用 NeuronChat,作者推断出包含从 VISp(或 ALM)细胞类型到所有交互对的目标区域的链接的通信网络,然后将它们聚合到所有交互对上。接下来,作者测试这个聚合网络是否可以预测通过逆行标记识别的投射网络。作者通过设置通信强度阈值(Fig. 2b, g)对聚合网络进行二值化,以计算灵敏度(NeuronChat 预测的链接与逆行标记所识别的链接的比例)和假阳性率(NeuronChat 预测的链接与未通过逆行标记识别的链接的比例)对于给定阈值。通过扫描一组连续阈值重复此过程,获得接收者操作特征(ROC)曲线。然后,使用 ROC 曲线下面积 (AUROC) 来衡量 NeuronChat 的预测性能:值越接近1,预测性能越好;随机分类器的 AUROC 值预计为 0.5。VIsp(Fig. 2c)和 ALM(Fig. 2h)的 AUROC 值分别为 0.832 和 0.764。请注意,对于这两种情况,预测的通信链路的一小部分都是不正确的(例如 3/21 for Fig. 2b and 7/30 for Fig. 2g)。作者还计算了另一个评估指标——精确率-召回率曲线下的面积(AUPRC),它总结了召回率(也称为灵敏度)和精度(通过逆行标记识别的链接与通过 NeuronChat 预测的链接的比例)之间的权衡,在不同阈值下。NeuronChat 预测的投射网络显示,VISp 的 AUPRC 为 0.915,ALM 的 AUPRC 为 0.768,明显优于随机分类器(Fig. 2d, i)。为了确定 ground truth 标签的特定图拓扑(即通过逆行标记识别的投射网络)是否使 NeuronChat 的预测任务变得容易,作者通过打乱图节点的细胞类型标签同时保持相同的图来扰动 ground truth 标签拓扑,然后重新计算 AUROC 和 AUPRC。作者发现,打乱后的 ground truth 标签的 AUROC 导致平均值约为 0.5 的分布,这表明即使具有相同的拓扑,这些打乱后的标签的预测能力也很差(Supplementary Fig. 1a, c, e, g)。此外,原始 ground truth 标签的 AUROC 显着高于打乱标签的 AUROC(VISp 和 ALM 投射网络的 p 值分别为 0.010±0.0036 和 0.017±0.0048)。AUPRC 的计算也得到了类似的结果(Supplementary Fig. 1b, d, f, h),这表明 NeuronChat 的预测能力并不能直接反映真实标签的特定图拓扑。
此外,AUROC 和 AUPRC 在输入数据二次采样率范围从 10% 到 90% 之间几乎没有变化(Supplementary Fig. 2)。虽然由于排列测试中的有限采样,预测的通信网络可能会在不同的重复模拟之间波动,但 NeuronChat 中使用的默认排列数量 (100) 会产生一致的 p 值,并且大多数重要链接是由 1000 个排列生成的(Supplementary Fig. 3)。综合起来,NeuonChat 对于输入数据的子采样和排列数量相对稳健。
作者还计算了各个交互对的通信网络的 AUROC 和 AUPRC(Fig. 2e, j 分别针对 VISp 和 ALM)。具有较高 AUROC 或 AUPRC 的交互对意味着更好的连接预测。正如预期的那样,各个交互对的 AUROC 和 AUPRC 的排名几乎相同,表明具有高 AUROC 的交互对通常表现出高 AUPRC。在与最高 AUROCs 的前 10 种相互作用中,VISp 的 7 种和 ALM 的 9 种是由谷氨酸介导的,这与投射神经元通常具有谷氨酸能,以及谷氨酸是大脑中主要的兴奋性神经递质这一事实相一致。例如,谷氨酸和 Grik5(glutamate receptor, ionotropic kainate 5)之间的相互作用,以及谷氨酸和 Gria2(AMPA 受体的亚基之一)之间的相互作用,在预测 VISp 和 ALM 预测的 AUROC 中排名前 3;谷氨酸与其他受体(例如 Gria1、Gria3、Grin1、Grin2b、Grm4、Grm7 和 Grik4)之间的相互作用也显示出 VIsp 或 ALM 的高 AUROC。此外,突触粘附分子 Neurexin 1 (Nrxn1) 和 Neuroligin 1 (Nlgn1) 之间的相互作用分别连接突触前和突触后神经元,在突触形成和成熟中发挥重要作用,在 AUROC 中排名第 10,在 AUPRC 中排名第 6,用于预测 ALM 投射。有趣的是,尽管 Neuroligin 2 被认为位于抑制性突触上,但 Nrxn2-Nlgn2 相互作用显示出对 VISp 投射的中等预测能力。令人惊讶的是,神经肽 Cck(胆囊收缩素)与其同源受体 Cckbr 之间的相互作用显示出对 VISp 投射的最高预测能力,表明 Cck-Cckbr 相互作用可能与长程神经元连接有关。神经肽 Penk 与其同源受体 Oprm1 之间的相互作用也显示出 VIsp 和 ALM 的高 AUROC。因此,当实验连接数据可用时,NeuonChat 能够发现与神经连接相关的具有生物学意义的交互对;当实验连接数据不可用时,具有高信息流的交互对(定义为所有检测到的重要链接的通信强度之和)可能是神经连接的潜在候选者,因为 AUROC(或 AUPRC)被发现具有中等的与信息流呈正相关(Supplementary Fig. 4)。
接下来作者用例子来说明 NeuronChat 如何利用发送细胞组中的配体和/或接收细胞组中的目标的丰度分布的变化来区分通信强度。虽然所有类型的谷氨酸能神经元都使用谷氨酸作为主要的神经递质,但配体丰度分布在 VISp 中的发送细胞组之间显示出明显的差异(Supplementary Fig. 5a,上图):大多数配体中 L2/3 IT、L4、L5 IT、L6 IT 比 L5 PT、L6 CT、L6b 更丰富。同样,目标丰度分布显示不同目标区域的接收细胞存在巨大多样性(Supplementary Fig. 5a, lower panel):主要目标在对侧 VIsp 中最丰富,而在 ACA 中最少。这些总体差异与以下事实一致:L2/3 IT、L4、L5 IT 和 L6 IT 比其他发送细胞组具有更多的传出链路,并且对侧 VISp 和 RSP 比 ACA 具有更多的传入链路(Fig. 2a, b)。在单个相互作用水平上,例如,对于 Glu-Gria2 相互作用对,L2/3 IT、L4、L5 IT 和 L6 IT 中与谷氨酸合成和运输相关的基因相对较高的表达使得这些细胞类型成为推断通信网络的主要发送者;对侧 VIsp 和 RSP 表达的 Gria2 高于 ACA,因此被推断为主要接收者(Supplementary Fig. 5b)。对于 Cck-Cckbr 相互作用对来说,这种模式更加清晰(Supplementary Fig. 5c)。还对 ALM 投射网络进行了分析,显示了类似的结果(Supplementary Fig. 6 )。总而言之,发送细胞组中的配体和/或接收细胞组中的目标的丰度分布的变化使得 NeuronChat 能够区分通信强度。
3. 与其他细胞间通讯推理工具和建模设置的比较
作者进一步将 NeuronChat 与两种流行的细胞间通信推理 (CCI) 工具 CellChat 和 CellPhoneDB 在使用相同 ligand-target 数据库(即 NeuronChatDB)预测神经元连接方面进行比较。作者使用与 NeuronChat 相同的计算流程来实现 CellChat 和 CellPhoneDB,除了配体丰度的计算和通信强度的公式(详细信息请参见比较方法)。在 VISp 和 ALM 投射网络中的神经元连接推理(参见上面的部分)中,NeuronChat 在两个方面优于现有的 CCI 方法:(1) 对于聚合通信网络,NeuronChat 在三种基准测试方法中具有最高的 AUROC 和 AUPRC(Fig. 3a, b for VISp and Fig. 3f, g for ALM); (2) 对于单个通信网络,NeuronChat 不仅检测到更多的交互对,而且比其他两种方法产生更高的 AUROC 和 AUPRC(Fig. 3c–e for VISp and Fig. 3h–j for ALM)。由于 CellChat 和 CellPhoneDB 对于相同功能的冗余基因都使用 AND 逻辑(几何平均值或最小值)而不是 OR 逻辑,因此小分子神经递质的丰度被大大低估,导致检测到的相互作用对比 NeuronChat 少。这些结果证明了 NeuronChat 在预测神经特异性细胞间通信方面的优势。

a. 三种方法的典型 ROC 曲线和 PR 曲线。
b. 三种方法推断的聚合 VISp 投射网络 100 次重复的 AUROC(左)和 AUPRC(右)值的箱线图。每个箱线图代表 100 个独立的重复计算。
c. 三种方法检测到的交互对的数量。
d-e. 三种方法推断出的单个 VISp 投射网络的 AUROC (d) 和 AUPRC (e) 值的箱线图。
f-j. 对 ALM 进行的相同分析。
虽然 NeuronChat 使用合成酶和囊泡转运蛋白的表达来估计小分子神经递质的丰度,但最近的方法 scFEA 使用图神经网络模型通过结合代谢和途径的化学计量效应来估计 scRNA-seq 的代谢通量和平衡依赖性。为了研究小分子神经递质的不同丰度替代物在识别神经特异性通讯网络中的影响,以谷氨酸为例,作者比较了 NeuronChat 的 ligand 丰度和八个 scFEA 衍生替代物(包括代谢物平衡和七个模块通量)在预测 VISp 和 ALM 投射网络中的效果。对于九个谷氨酸替代物中的每一个,计算了 24 个谷氨酸介导的相互作用对的通信网络的 AUROC 和 AUPRC 值,发现 NeuronChat 的配体丰度在九个谷氨酸替代物中的 AUROC(或 AUPRC)中值中显示出中等或以上的排名(Supplementary Fig. 7a, c)。对于聚合超过 24 个谷氨酸介导的相互作用对的通信网络,NeuronChat 的配体丰度在 AUROC 和 AUPRC 中均排名第二,用于预测九个谷氨酸替代物中的 VIsp 和 ALM 投射网络(Supplementary Fig. 7b, d)。尽管如此,NeuronChat 的配体丰度与最好的 scFEA 衍生替代物之间的差异非常小。这些结果表明,NeuronChat 的配体丰度尽管简单,但在神经元连接的推断上效果良好。
NeuronChat 使用 Tukey's trimean 而不是算术平均值来计算组中所有细胞中基因表达的平均值。根据他们的定义,NeuronChat 使用 Tukey's trimean 而不是算术平均值来计算组中所有细胞中基因表达的平均值。根据他们的定义,非零 Tukey's trimean 仅在该基因在至少 25% 的细胞中表达时出现,而非零算术平均值在该基因在至少一个细胞中表达时出现,Tukey's trimean 有利于识别细胞类型富集的 ligand–target 对。正如预期的那样,Tukey's trimean 导致检测到的相互作用对少于算术平均值(Supplementary Fig. 8a, e);然而,Tukey's trimean 产生的相互作用对总体上显示出比算术平均产生的相互作用对更高的 AUROC 和 AUPRC,这表明 Tukey's trimean 能够推断出更可靠的相互作用对(Supplementary Fig. 8b–d and 8f–h)。
作者还研究了四种不同聚合方法对推断神经元连接的影响。对于“thresholded weight”方法,选择交互对的阈值作为交互对所有通信强度值的 80% 分位数(基准测试的默认设置)。这是因为 80% 分位数导致整体 AUROC/AUPRC 高于其他阈值分位数,但 ALM 投射网络的 AUROC 略低(Supplementary Fig. 9a, b and 9e, f)。在四种聚合方法中,作者发现“thresholded weight”方法为 VIsp 投射网络产生最高的 AUROC/AUPRC 值,而第二高的 AUROC/AUPRC 值仅略低于 ALM 投射网络的最佳值(Supplementary Fig. 9c, d and 9g, h)。此外,与重复模拟的其他聚合方法相比,“thresholded weight”导致 AUROC/AUPRC 的变化更小,从而稳健地最小化排列测试中产生的随机性。
4. NeuronChat 识别 VISp 中的细胞间通讯模式和功能相关的相互作用
NeuronChat 的基本功能是推断和可视化细胞间通信网络,然后识别细胞间通信模式并对功能相关的交互对进行分类。作者通过将 NeuronChat 应用于小鼠 VISp 的 scRNA-seq 数据来说明 NeuronChat 的这些功能。15,469 个单细胞被很好地注释为 3 个细胞类别(Glutamatergic, GABAergic, and Non-Neuronal)和 21 个细胞亚类。通过将 NeuronChat 应用于该数据集,识别了 21 个细胞亚类之间的通信网络,可以通过圆形图、弦图和热图进行可视化(Fig. 4a)。每个谷氨酸能亚类向大多数 GABA 能亚类和所有谷氨酸能亚类以及星形胶质细胞发送信号,而这些链接的通信强度可能不同。有趣的是,谷氨酸能亚类之间存在密集的交流。与谷氨酸能亚类相比,GABA 能亚类与三种细胞类的传出通讯相对稀疏。GABA 能亚类中,Lamp5、Sncg 和 Vip 是主要发送者;Lamp5、Pvalb 和 Vip 亚类接收来自 GABA 能亚类和谷氨酸能亚类的信号,而 Sst 和 Meis2 亚类则显示出偏好接收来自谷氨酸能神经元的信号。
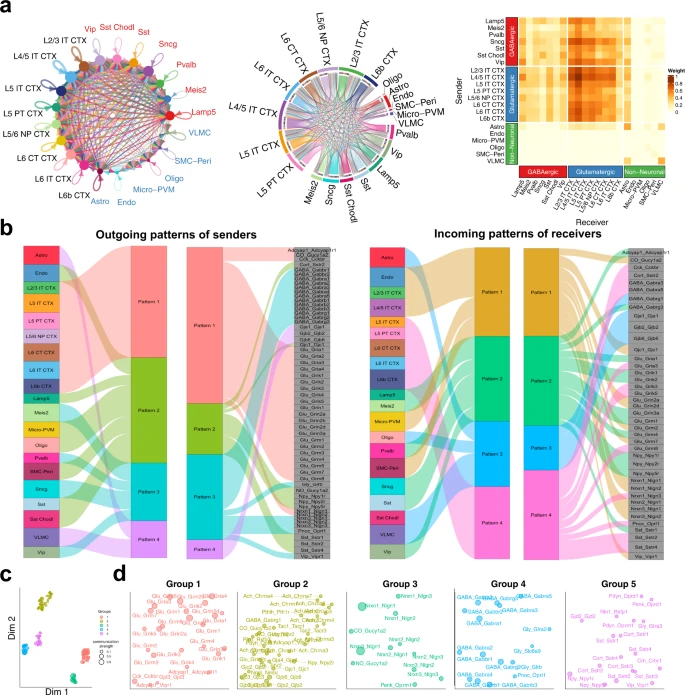
a. 对 VISp 的多种细胞类型之间推断出的通信网络的可视化。圆形图、弦图和热图用于可视化通过 "weight" 方法聚合的细胞间通信网络。在 circle plot 中,节点大小表示细胞发出信号的强度,文本标签用不同颜色标记,以表示它们的细胞类别(即 glutamatergic, GABAergic and non-neuronal);每个连接的通信强度由宽度表示。在 chord diagram 中,不同细胞类别的扇形之间有较大的间隙;每个连接的宽度表示通信强度,而扇形的宽度(代表一个细胞群体)反映了来自或到该细胞群体的总通信强度。在 heatmap 中,通过颜色条表示通信强度。
b. 发送者的发出信号模式和接收者的接收信号模式通过冲积图可视化,显示推断出的潜在模式与细胞群体之间的对应关系,以及推断出的潜在模式与相互作用对之间的对应关系。流的厚度表示细胞群体或相互作用对对每个潜在模式的贡献。每个模式的高度与其相关联的细胞群体或相互作用对的数量成比例。用于分析的是信息流最高的前60个相互作用对。
c. 根据它们的功能相似性将相互作用投影到二维流形上。每个相互作用对由一个点表示,其颜色和大小分别表示功能组和总通信强度(按最大值归一化)。
d. 每个相互作用组的二维流形放大视图。
总共检测到 109 个重要的交互对以及所有交互对的链接数量(Supplementary Fig. 10)。计算每个相互作用对的信息流后,相互作用对 Nrxn1-Nlgn1、Nrxn3-Nlgn1、Glu-Gria2、Glu-Grin2b、GABA-Gabra1、Glu-Gria4、GABA-Gabrb1、Glu-Grm5、GABA-Gabrg2 和 Glu -Grik2 在信息流中排名前 10,而 NO-Gucy1a2、Glu-Gria2、Glu-Grin1、Glu-Grin2b、GABA-Gabbr1、GABA-Gabrb1、Nrxn1-Nlgn1、GABA-Gabrb3、Glu-Gria1 和 Nrxn3- Nlgn1 的链接数排名前 10。信息流中的前 10 名与链接数量中的前 10 名之间的不完全重叠表明某些相互作用(例如 Glu-Gria4)具有很强的个体链接,而其他一些相互作用(例如 Glu-Grin1)则显示出特定的相互作用。细胞亚类之间的广泛通信,但单个链接的强度中等。
使用模式识别方法,NeuronChat 检测发送细胞的传出模式和接收细胞的传入模式,这可以通过冲积图可视化。这使得发送/接收细胞类型和潜在模式之间的对应关系以及潜在模式和单个相互作用对之间的对应关系可视化(Fig. 4b)。正如预期的那样,所有谷氨酸能亚类都对应于输出模式 #1,该模式与所有谷氨酸信号传导、Nrxn1 信号传导和一些神经肽相互作用(例如 Cck-Cckbr 和 Adcyap1-Adcyap1r1)相关。与抑制性神经元的多样性一致,GABA 能亚类对应于输出模式 #2-3:Lamp5、Sst 和 Sst Chodl 亚类属于模式 #2,主要与神经肽 Sst、Npy 和 Cort 的信号传导有关;Pvalb、Meis2、Sncg 和 Vip 亚类属于模式 #3,其中包括 GABA、甘氨酸和 Nrxn3 以及一些神经肽(例如 Vip 和 Pnoc)的信号。对于非神经元亚类,Endo、Micro-PVM 和 SMC-Peri 对应于传出模式 #2,而 Astro 和 VLMC 属于代表间隙连接蛋白相互作用的模式 #4。与传出模式不同,传入模式 #2 和 #4 同时包含谷氨酸信号和 GABA 信号,表明相应的细胞亚类(例如 L5 IT CTX 和 Lamp5)同时接收兴奋性和抑制性输入。然而,谷氨酸能亚类和 GABA 能亚类对应于不同的传入模式(分别为模式 #1 和 #4,以及模式 #2-3)。
通过流形学习,NeuronChat 将交互对投影到二维流形中,并根据通信网络的功能相似性将它们分组到不同的簇中。功能相似性衡量交互对共享相似发送者和接收者的程度。109 个交互对被分为 5 个独立的组(Fig. 4c、d)。Group #1 以谷氨酸信号为主,代表来自谷氨酸能亚类的信号传导,而 Group #4 包括许多 GABA 和甘氨酸信号,代表来自 GABA 能亚类的信号传导(另请参见 Supplementary Fig. 11 中每个相互作用组的聚合通信网络)。神经毒素和神经连接蛋白之间的相互作用在 Group #3 中占主导地位。Group #5 包含神经肽信号传导,例如来自 GABA 能亚类的 Sst、Vip、Penk 和 Npy,表明这些相互作用对具有相似的通信模式。Group #2 以间隙连接蛋白和一些神经肽信号为主,主要代表非神经元亚类之间的信号传导。流形学习的结果表明,NeuronChat 能够将检测到的 ligand-target 相互作用对分类为具有生物学意义的组。
5. NeuronChat 揭示了 ALM 和 VISp 层间兴奋网络之间保守且特定于上下文的通信模式
NeuronChat 的另一个应用是对不同的生物环境进行比较,以识别保守的或特定于环境的通信模式。在这里,作者通过比较 ALM 和 VISp 之间的层间兴奋性通信网络来说明 NeuronChat 的此类功能。scRNA-seq 数据来自已发表的论文,包括 ALM 的 4,600 个谷氨酸能神经元和 VIsp 的 8,114 个谷氨酸能神经元,两者都分为 8 个共享子类。虽然两者的总体通信模式相似(Fig. 5a),但各个交互对的一些通信网络是不同的。例如,ALM 和 VISp 的 Glu-Grin3a 通信网络显示出显着差异(Fig. 5b):L4/5 IT CTX 预计是 ALM 的主要靶细胞亚类之一,但不是 VISp 的主要靶细胞亚类之一。
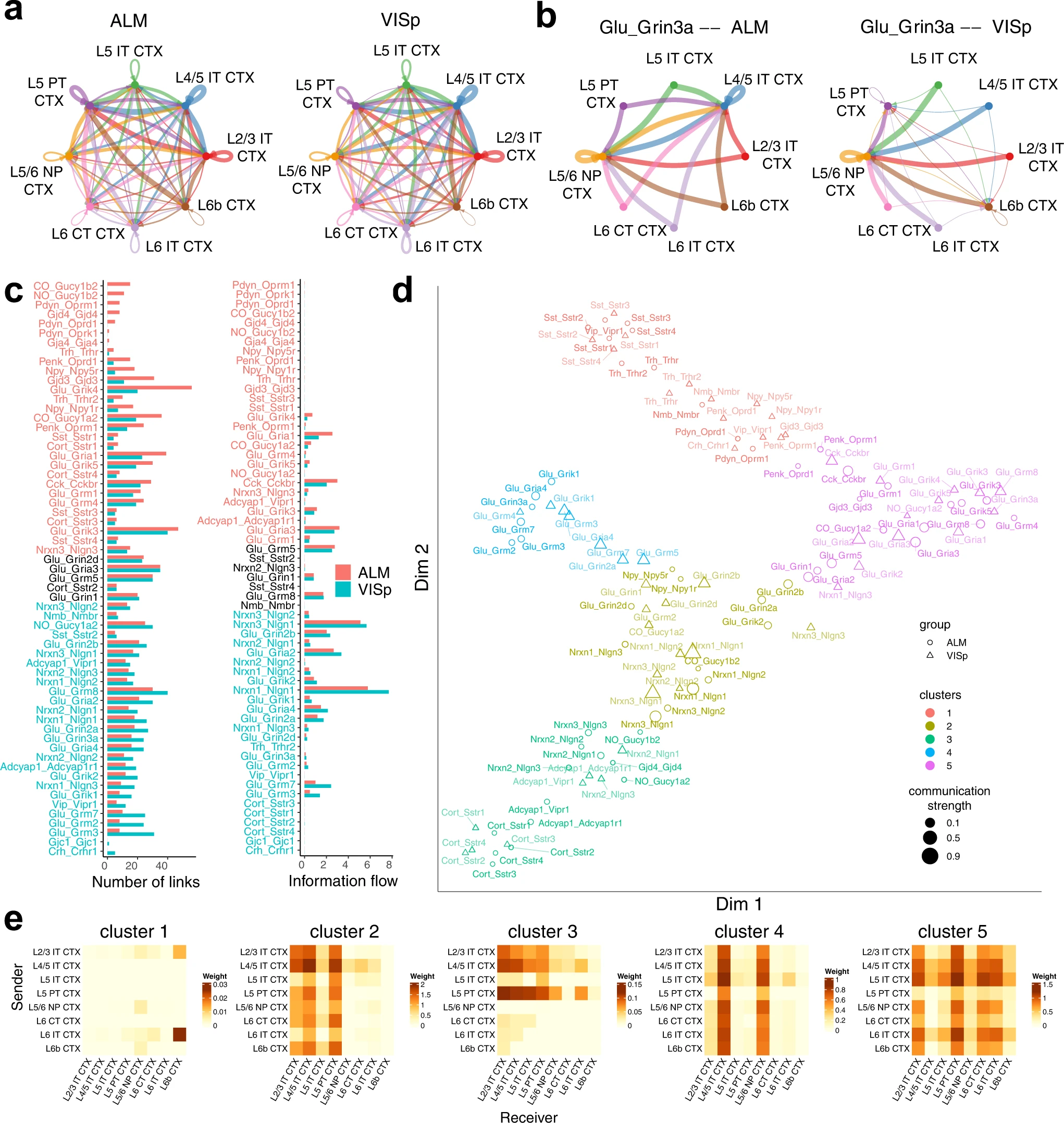
a. ALM(左侧)和 VISp(右侧)的聚合层间兴奋性通信网络。使用的聚合方法是 "weight" 。
b. Glu-Grin3a 的 ALM(左侧)和 VISp(右侧)的单个层间兴奋性通信网络。
c. 柱状图比较 ALM 和 VISp 之间每个相互作用对的连接数(左侧)和信息流(右侧)。
d. 将 ALM 和 VISp 的相互作用投影到二维流形上,根据它们的功能相似性。每个相互作用对由一个圆形(对于VISp)或三角形(对于ALM)表示,其颜色和大小分别表示功能组和总通信强度(按最大值归一化)。
e. (d)中五个相互作用簇的聚合层间兴奋性通信网络的热图。
为了确定单个通信网络的差异,作者比较了两个区域之间所有单个交互的链接数量和信息流(Fig. 5c)。发现只有少数相互作用对在 ALM 和 VISp 之间共享相同或接近相同的信息流(或链接数量),例如 Glu-Grin1 和 Glu-Grm5。其他相互作用对显示两个区域之间存在显着差异,例如,Pdyn 信号传导(包括 Pdyn-Oprm1、Pdyn-Oprk1 和 Pdyn-Oprd1)在 ALM 中完全开启;Glu-Grik4 和 Glu-Gria1 通信网络在 ALM 中比在 VIsp 中密集得多;与 ALM 相比,VISp 中的 Glu-Grm3 和 Glu-Grm7 通信网络以及由 Nlgn1 介导的通信网络显示出更多的链接和更高的信息流。
比较 ALM 和 VISp 各个通信网络的另一种方法是基于它们的功能相似性。通过根据功能相似性将 VISp 和 ALM 的相互作用对投影到相同的二维流形上,然后将它们分类为簇,我们可以进一步发现保守的或特定于上下文的相互作用对(Fig. 5d)。如果一个交互对的通信模式在 ALM 和 VISp 之间是保守的,则 ALM 和 VISp 的此类交互对的通信网络应分组到同一集群中,反之亦然。二维流形显示所有配体-靶标相互作用对都分为五个簇,每个簇的聚合网络由热图显示(Fig. 5e):cluster #1,与神经肽信号传导(例如,Sst、Trh、Nmb、Pdyn 和 Vip)相关,代表从 L6 IT CTX 到 L6b CTX 的信号主导的稀疏通信模式;cluster #2,包括谷氨酸信号传导和神经素-神经配素相互作用,显示了 L2/3 IT CTX、L4/5 IT CTX 和 L5 PT CTX 是主要接收者的通信模式;cluster #3 代表通信模式,其中信号主要来自 L2/3 IT CTX、L4/5 IT CTX 和 L5 PT CTX;cluster #4 显示了清晰的模式,其中 L4/5 IT CTX 和 L5/6 NP CTX 接收来自所有子类的信号;cluster #5 代表子类之间密集且强大的通信,主要由谷氨酸信号主导。有趣的是,对于大多数 ligand-target 相互作用对,ALM 和 VISp 的通信网络被分类为相同的簇,表明大多数 ligand-target 相互作用对在两个区域之间可能是保守的。例如,ALM 和 VISp 中 Glu-Gria2 的通信网络都被分组到 cluster #5 中。然而,用于 ALM 和 VISp 的 Glu-Grin3a 通信网络分别分为cluster #4 和 cluster #5,这表明 Glu-Grin3a 对于 ALM 和 VISp 是上下文特定的(see also Fig. 5b);同样,用于 ALM 和 VISp 的 Nrxn1-Nlgn3 的通信网络分别属于 cluster #2 和 cluster #5。
6. NeuronChat 预测自闭症谱系障碍患者细胞间通讯模式的变化
NeuronChatDB 还包含人类的 ligand-target 相互作用信息。作者通过将 NeuronChat 应用于已发表的自闭症谱系障碍 (ASD) 患者和健康对照的皮质组织单核 RNA 测序数据,研究如何使用 NeuronChat 来预测特定神经系统疾病患者细胞间通讯模式的变化。数据收集自 15 名 ASD 患者和 16 名对照者的死后组织样本,包括前额皮质和前扣带皮层,其中包含 ASD 患者的 52,003 个单核和对照的 52,556 个单核。为了降低计算成本,总共采样了 20,000 个细胞进行分析(10,000 个用于 ASD,10,000 个用于对照)。
作者使用 NeuronChat 推断 ASD 患者和对照组 17 种细胞类型之间的通信网络,包括兴奋性神经元、中间神经元和星形胶质细胞的亚型。对于 NeuronChat 推断的通信网络,ASD 不仅显示出比对照更多的总链接(Fig. 6a,上图),而且还增加了链接强度(Fig. 6a,下图),表明 ASD 中细胞类型之间的通信总体增强。与此一致的是,通过对比 ASD 和对照组的聚合细胞间通讯网络,发现与对照组相比,ASD 患者的细胞间通讯上调(Fig. 6b,上图)多于下调(Fig. 6b,下图)。这可能支持自闭症局部过度连接的假设。此外,对于每种细胞类型和每个相互作用对,作者计算了 ASD 中与对照组相比的差异传出和传入通信强度(Fig. 6c)。从差异传出模式(Fig. 6c,上图)来看,观察到大多数兴奋性神经元类型的大多数谷氨酸信号都增强了;对于大多数细胞类型,由 Neuroligin 1 介导的传出信号得到增强,但在某些细胞类型(如 PV 中间神经元、少突胶质细胞或 OPC(少突胶质细胞前体细胞))中受损。从传入的差异模式(Fig. 6c,下图)中,观察到,虽然向大多数细胞类型发出的谷氨酸信号增强,但向少突胶质细胞(NRXN1-NLGN1 和 NRXN3-NLGN1)发出的 Neuroligin 1 信号以及向 OPC 发出的 Neuroligin 3 信号( NRXN1-NLGN3、NRXN2-NLGN3 和 NRXN3-NLGN3) 大大减少。有趣的是,NLGN3 基因位点中超过 10 个错义突变已被确定与 ASD 相关,并且在 Nlgn3 突变的动物模型中出现了与 ASD 相关的行为表型(例如异常的社交互动、刻板行为和增强的空间学习)。作者的结果表明,NLGN3 信号传导的下调也可能是 ASD 功能障碍的基础。事实上,ASD 动物模型显示少突胶质细胞数量和髓鞘形成减少,而 OPC 向少突胶质细胞的分化则受到 NLGN3 的影响。与这一证据一致,作者的分析中发现,自闭症谱系障碍 (ASD) 中 NLGN3 信号传导至 OPC 的特异性下调表明,有缺陷的 NLGN3 信号传导可能通过 OPC 功能障碍的机制导致自闭症谱系障碍 (ASD)。
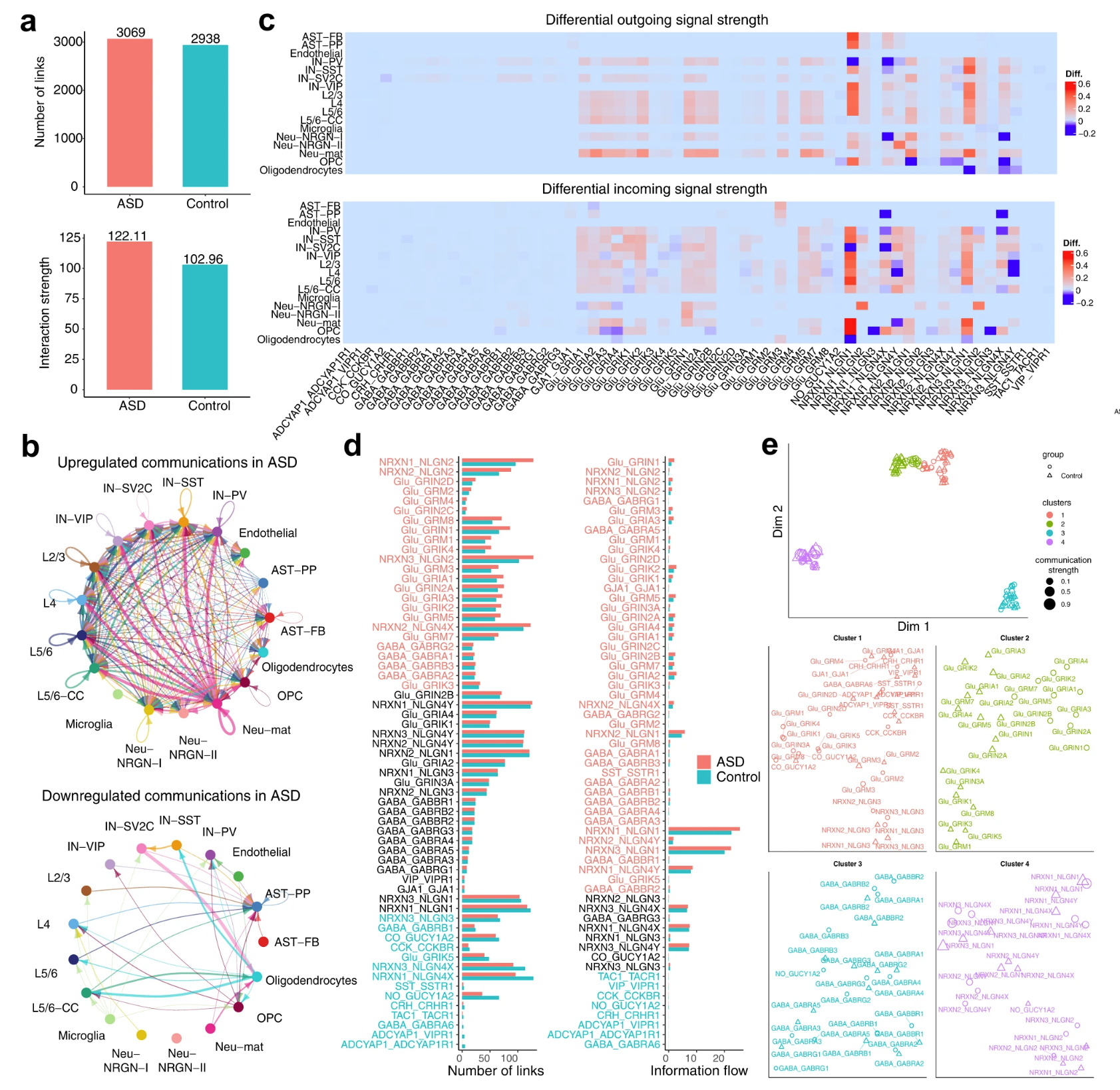
a. 比较 ASD 和对照组之间推断出的所有连接的数量(上)和权重(下)。
b. 圆形图显示与对照组相比,ASD中上调(上)和下调(下)的细胞间通信。每个连接的宽度表示 ASD 和对照组之间所有相互作用对的通信强度值之和的绝对差值。
c. 热图显示 ASD 和对照组之间输出(上)和输入(下)信号强度的差异。给定细胞群体和相互作用对,输出(或输入)信号强度定义为来自(或到)细胞群体的连接的通信强度之和。颜色条表示 ASD 和对照组之间输出(或输入)信号强度值的差异。
d. 柱状图比较 ASD 和对照组之间每个相互作用对的连接数(左侧)和信息流(右侧)。
e. 根据它们的功能相似性将相互作用投影到二维流形上(上部),并放大每个相互作用组(下部)。每个相互作用对由一个圆形(对于ASD)或三角形(对于对照组)表示,其颜色和大小分别表示功能组和总通信强度(按最大值归一化)。
接下来,作者比较 ASD 和对照之间每个交互对的通信链路数量和信息流(Fig. 6d)。ASD 和对照共享相似数量的链接和相似的交互对信息流,例如 GABA-GABRG3 和 NRXN1-NLGN4Y,这可能不是 ASD 所特有的。一些相互作用对,如 NRXN2-NLGN2、Glu-GRM3 和 Glu-GRIN1,在 ASD 中的链接数量和信息流均上调。许多相互作用对在链接数量上表现出差异不大,但信息流增加,例如 NRXN2-NLGN1、Glu-GRIA2 和 Glu-GRIA4。其他相互作用,如 CCK-CCKBR、CRH-CRHR1、TAC1-TACR1 和 GABA-GABRA6,在链接数量和信息流方面都显示 ASD 减少。
作者通过将 ASD 和对照的相互作用对投影到相同的二维流形上并根据其功能相似性进行聚类,进一步识别保守的或特定于上下文的相互作用对(Fig. 6e)。所有 ligand-target 相互作用对分为四个 cluster:cluster 3 和 cluster 4 分别由 GABA 信号传导和神经毒素-神经配蛋白信号传导主导;cluster 1 和 cluster 2 均以谷氨酸信号传导为主,而 cluster 1 还包括神经肽信号传导和间隙连接相互作用以及 Neuroligin 3 信号传导。令人惊讶的是,对于大多数交互,自闭症谱系障碍和对照的通信网络被分组到相同的 cluster 中,这表明自闭症谱系障碍和对照之间的大多数通信模式在功能相似性方面是保守的。对于交互对 Glu-GRIK1、Glu-GRIK3、Glu-GRIK4、Glu-GRIK5 和 Glu-GRIN3A,ASD 和控制的通信网络分别分为 cluster 1 和 cluster 2。这意味着这些交互对的发送者或接收者在 ASD 和对照之间是不同的。
7. NeuronChat 利用空间转录组学来推断和可视化神经特异性通信网络
NeuronChat 还可用于根据空间转录组学 (ST) 数据推断和可视化神经特异性通信网络,这些数据测量神经细胞中的基因表达及其空间位置。作者通过将 NeuronChat 应用于基于三种不同测序技术(包括 seqFISH+、MERFISH 和 Visium)的三个小鼠大脑 ST 数据集来说明 NeuronChat 的此类功能。seqFISH+ 数据集包括小鼠体感皮层和室下区 913 个细胞中 10,000 个基因的 mRNA 表达,其中有 4 种类型的 358 个兴奋性神经元。MERFISH 数据集包括小鼠初级运动皮层和邻近区域约 300,000 个细胞(包括 9 个谷氨酸能亚类和 5 个 GABA 能亚类以及非神经元亚类)中 258 个基因的 mRNA 表达。Visium 数据集包括小鼠大脑冠状切片 2,702 个点的 mRNA 表达谱,这些点被分为七个簇。
对于所有三个 ST 数据集,作者计算细胞类型(或点簇)之间的通信网络,而无需施加空间限制。对于 seqFISH+ 和 Visium 数据集,通信网络直接根据空间转录组计算;对于 MERFISH 数据集,由于 MERFISH 数据集中包含的基因数量太少,无法覆盖大部分 ligand-target 对,因此使用小鼠初级运动皮层的 scRNA-seq 数据来推断七种兴奋性细胞类型之间的通信网络由 MERFISH 数据集和 scRNA-seq 数据集(7 个谷氨酸能亚类的 4,461 个细胞)共享。为了可视化空间中的通信网络,作者开发了一种多层可视化工具来共同说明空间通信网络、细胞类型/点簇注释和组织图像/解剖参考(Fig. 7a–c; Supplementary Fig. 12)。
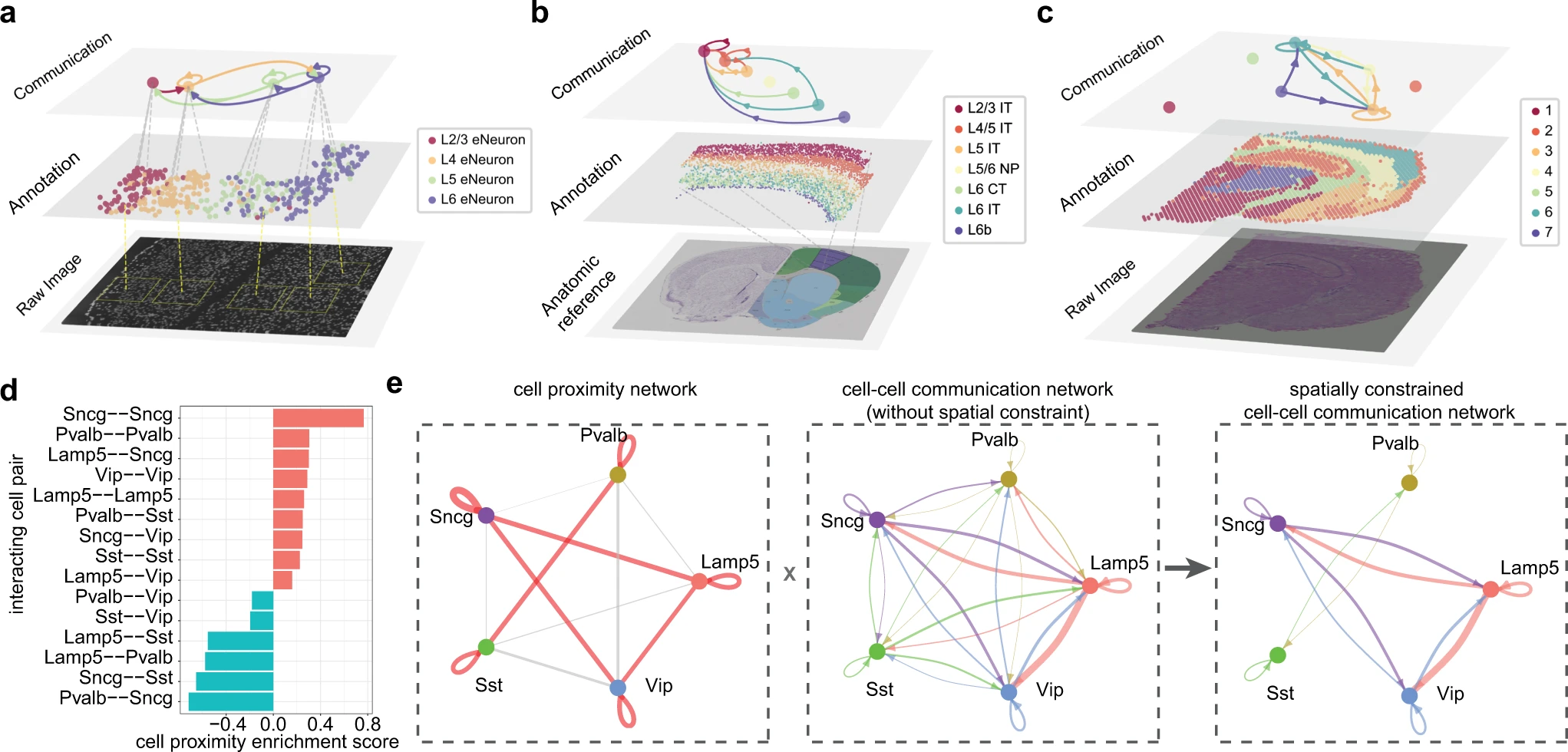
a-c. 由不同技术生成的三个空间转录组学数据集的多层可视化,包括seqFISH+(a)、MERFISH(b)和Visium(c)。每个图包括原始组织切片图像/解剖学参考(底部)、空间中的细胞/斑点注释(中间)和聚合通信网络,显示前10个连接(顶部)。连接的宽度表示所有显著配体-靶标对的通信强度之和。(b)底部图像是 Allen 大脑解剖学参考。
d. 柱状图显示所有成对相互作用细胞类型的细胞临近富集得分。细胞临近富集得分是基于所有 64 个 MERFISH 切片计算的。score>0(红色条)和score<0(青色条)分别表示相互作用细胞类型之间丰富和稀疏的临近。
e. GABA 能神经元空间约束通信网络的推断。左侧:细胞临近网络。红色或灰色链接分别表示相互作用细胞类型之间丰富或稀疏的临近;链接的宽度表示丰富或稀疏的强度。中间:无空间约束的细胞-细胞通信网络,基于scRNA-seq 数据计算,针对同一大脑区域和同一细胞类型(共2044个单细胞)。链接的宽度表示所有显著配体-靶标对的通信强度之和。右侧:空间约束的细胞-细胞通信网络,通过从原始细胞-细胞通信网络中删除具有稀疏临近性的链接获得。
NeuronChat 还提供了向通信网络添加空间约束的选项。作者基于 MERFISH(Supplementary Fig. 13)和 scRNA-seq 数据,通过将其应用于 MOp 皮层的五个 GABAergic 子类(即 Lamp5、Sncg、Vip、Sst 和 Pvalb)之间的通信来测试此功能。虽然神经特异性信号经常发生长距离空间通信,但 GABA 能神经元通常具有局部轴突乔木,并且它们之间的连接概率随着神经元间距离的增加而降低。为了研究 GABA 能神经元对通信网络的潜在空间影响,我们通过计算与之前的研究类似的空间邻近度富集分数来表征 GABA 能神经元之间的空间邻近度(Fig. 7d)。然后,我们可以删除空间邻近度得分低于给定阈值的通信链接,从而形成空间受限的通信网络(Fig. 7e)。例如,虽然 Pvalb 子类与通信网络中其他四个 GABA 能子类中的每一个都有连接而没有空间限制,但该子类仅显示出与其自身或 Sst 子类丰富的细胞接近度。这一观察结果与 Pvalb 细胞更愿意与其他 Pvalb 细胞连接的证据一致。
讨论
NeuronChat 专为从单细胞表达数据和空间解析转录组学推断神经特异性细胞间通讯而设计。作者构建了神经信号 ligand-target 相互作用数据库,并提出了一个计算模型,该模型结合了神经信号传输过程来推断细胞间通信,这使得 NeuronChat 不同于现有的用于推断细胞间通信的非神经元活动方法。NeuronChat 在多个数据集上的基准测试和应用显示了其预测神经连接的能力。考虑到从越来越多的单细胞转录组数据集和空间转录组数据集中识别出的神经元异质性,NeuronChat 可以预测不同转录组状态之间的新神经连接。通过将可用的神经连接数据(例如逆行标记数据和电生理学数据)与我们预测的特定配体-靶标相互作用对的通信网络(例如 Fig. 2e, j)进行对比,我们可以进一步识别可能的信号通路和关键基因。提供对神经连接背后的未表征机制的见解。
NeuronChat R 软件包提供了多功能且易于使用的可视化工具和网络分析方法,可以方便地探索神经特定的细胞间通信模式。在本研究中使用此类分析工具,NeuronChat 可以根据其传出信号模式将谷氨酸能神经元亚型和 GABA 能神经元亚型分类为单独的发送者组;由甘氨酸介导的相互作用对被分类为包含来自 GABA 能神经元的 GABA 信号的相互作用组;自闭症谱系障碍 (ASD) 患者中已发现 NLGN3 信号转导至 OPC 的特异性下调。总的来说,NeuronChat 能够通过 scRNA-seq 数据中具有生物学意义的发现来破译复杂的神经元间通信。为了探索预测的神经元间通讯下游的信号通路和基因调控网络,可以使用现有的数据库,如OmniPath、STRING 和 NicheNet,通过连接信号传导和转录调控来构建整合的细胞间通讯网络。
基于排列的 p 值计算的准确性和计算速度之间存在权衡,这是 NeuronChat 的所有新应用程序所需要的。大量排列可以产生更准确的经验 p 值,但计算时间较长。尽管如此,作者发现 100 和 1000 个排列计算出的 p 值之间具有良好的一致性,并且 1,000 个排列检测到的链接与 100 个排列检测到的链接在很大程度上重叠,丢失的链接不超过 1.5%(Supplementary Fig. 3)。这些结果表明,可以减少排列数量(例如 NeuronChat 中的默认数量 100),以节省计算时间,同时保持 p 值计算的准确性。
虽然 NeuronChat 的计算工作流程已经过优化以预测神经元连接性,但可以扩展设置以纳入更精细的模型,例如用于估计小分子神经递质的丰度。对于这种情况,除了仅使用合成酶和囊泡转运蛋白的表达之外,还可以包括代谢和途径依赖性的化学计量效应。通过比较九个谷氨酸丰度替代物,我们发现一些 scFEA 衍生的替代物显示出比 NeuronChat 的配体丰度更高的 AUROC(或 AUPRC)值(Supplementary Fig. 7)。虽然 NeuronChat 的配体丰度与最佳 scFEA 衍生替代物之间的差异很小,但它提出了提高神经元连接预测准确性的方法。
与其他推断细胞间通讯的现有方法一样,NeuonChat 估计转录组学中配体和靶蛋白的丰度,这些丰度可能与蛋白质或代谢物水平不一致。原则上,NeuronChat 可以应用于蛋白质组学和代谢组学数据,以推断配体-靶标相互作用(如果数据可用)。由于单细胞蛋白质组学和代谢组学技术在分子覆盖范围或通量上落后于转录组学,目前转录组学数据仍然是细胞间通讯推断的主要数据来源。
虽然 NeuronChatDB 包括主要的小分子神经递质、大部分神经肽、一些气体递质、间隙连接蛋白以及突触粘附分子,但在策划的交互条目中可能存在缺失信息,导致推理出现偏差。尽管如此,NeuronChat 允许使用当前版本中未包含的用户定义的交互轻松更新数据库,以扩展其更多交互的适用性。
在当前的研究中,NeuronChat 在评估其丰度时将异聚受体的多个不同亚基分成单独的条目。原则上,需要考虑亚基化学计量以更准确地代表异聚受体。异聚受体可以由各种亚基组合组装而成,亚基组成具有很大的多样性,极大地影响其功能特性。例如,AMPA 受体中 GluA2 亚基的存在决定了钙离子的渗透性。虽然 NeuronChatDB 不包含大多数异聚受体很大程度上未知的亚基化学计量信息,但 NeuronChat 包含使用客户提供的亚基化学计量对特定异聚受体丰度进行建模的选项。我们期望结合辅助蛋白和下游基因的知识将提高通信预测的准确性。
注:本文为个人学习笔记,仅供大家参考学习,不得用于任何商业目的。如有侵权,请联系作者删除。

本文由 mdnice 多平台发布